Общие сведения о цитокинах
При изучении дифференцировки иммунокомпетентных клеток и механизмов межклеточного взаимодействия при формировании как клеточного, так и гуморального иммунного ответа была открыта большая группа медиаторов белковой природы, названных в последующем цитокинами.
Основная биологическая активность цитокинов — регуляция иммунного ответа на всех этапах его развития: пролиферация и дифференцировка предшественников функционально активных иммунокомпетентных клеток, переработка и презентация антигена в иммуногенной форме на поверхности фагоцитирующих мононуклеаров, пролиферация антигенчувствительных лимфоцитов, дифференцировка В-клеток в продуценты иммуноглобулинов, переключение синтеза иммуноглобулинов с одного изотипа на другой, обеспечение созревания предшественников цитотоксических Т-клеток до зрелых эффекторов, индукция цитотоксичности у макрофагов. Некоторые из цитокинов являются активными участниками воспалительной реакции и острофазного ответа, проявляющих себя в элиминации опухолевых клеток.
Группы цитокинов:
- Интерлейкины (ИЛ).
- Факторы некроза опухоли.
- Интерфероны.
- Колониестимулирующие факторы (КСФ).
- Трансформирующий фактор роста-β (ТФР-β).
- Хемокины.
Механизмы действия цитокинов
Интракринный механизм – действие цитокинов внутри клетки-продуцента; связывание цитокинов со специфическими внутриклеточными рецепторами.
Аутокринный механизм – действие секретируемого цитокина на саму секретирующую клетку. Например, интерлейкины-1, -6 -18, ФНОα являются аутокринными активирующими факторами для моноцитов/макрофагов.
Паракринный механизм – действие цитокинов на близкорасположенные клетки и ткани. Например, ИЛ-1, -6 -12 и -18, ФНОα, продуцируемые макрофагом, активируют Т-хелпер (Th0), распознающий антиген и МНС макрофага (Схема аутокринно-паракринной регуляции иммунного ответа).
Эндокринный механизм – действие цитокинов на расстоянии от клеток-продуцентов. Например, ИЛ-1, -6 и ФНОα, помимо ауто- и паракринных воздеиствий могут оказывать дистантное иммунорегуляторное действие, пирогенный эффект индукцию выработки белков острой фазы гепатоцитами, симптомы интоксикации и мультиорганные поражения при токсико-септических состояниях.
Аутокринно-паракринная регуляция иммунного ответа
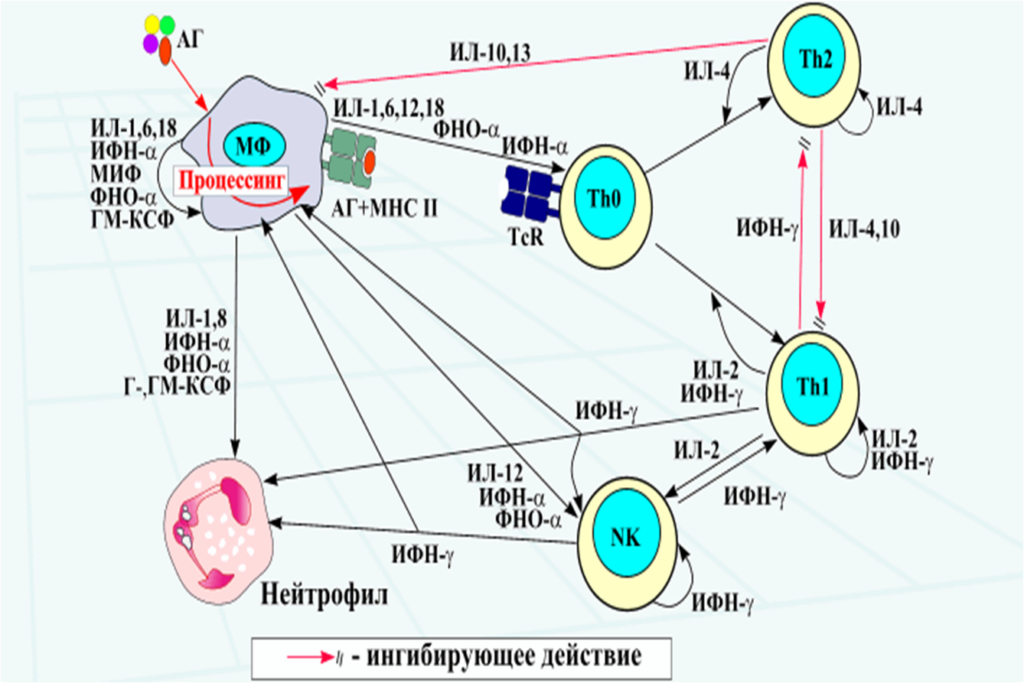
Интерлейкины – цитокины, ответственные за межклеточные взаимодействия между лейкоцитами Описано около 20 интерлейкинов.
Ил-1 продуцируется гл. обр. макрофагами и в меньшей степени дендритными клетками, эндотелиоцитами, фибробластами, NK, кератиноцитами, некоторыми клонами Th2. Он стимулирует продукцию Т-хелперами ИЛ-2, способствует проявлению рецепторов к ИЛ-2 на Т-лимфоцитах, влияет на созревание В-лимфоцитов, а также оказывает провоспалительное и пирогенное действие. Стимулирует образование гепатоцитами белков острой фазы, усиливает функции нейтрофилов, NK, обеспечивает взаимосвязь иммунной, нервной и эндокринной систем. Оказывает провоспалительное и пирогенное действие, обеспечивает взаимосвязь иммунной, нервной и эндокринной систем.
Ил-2 вырабатывается Т-лимфоцитами, а также цитотоксическими лимфоцитами (CD8+) 1 порядка. Он активирует дифференцировку Thl и Т-киллеров, стимулирует NK и синтез иммуноглобулинов В-лимфоцитами.
Ил-3 продуцируется Т-лимфоцитами и стволовыми клетками. Является ростовым фактором стволовых и ранних предшественников гемопоэтических клеток.
Ил-4 продуцируется гл. обр. Th2. Он стимулирует дифференцировку Th0 в Th2, стимулирует синтез иммуноглобулинов В-лимфоцитами, подавляет генерацию цитотоксических лимфоцитов, NK, а также продукцию ИФН-g и противоопухолевую активность макрофагов.
Ил-5 синтезируется Th2. Способстует пролиферации и дифференцировке стимулированных В-лимфоцитов, усиливает продукцию IgA, активирует эозинофилы.
Ил-6 вырабатывается макрофагами, Т- и В-лимфоцитами. Стимулирует пролиферацию тимоцитов, В-лимфоцитов, активирует предшественников цитотоксических лимфоцитов, гранулоцитов и макрофагов, стимулирует образование гепатоцитами белков острой фазы, оказывает провоспалительное действие, обеспечивает взаимосвязь иммунной, нервной и эндокринной систем.
Ил-7 продуцируется стромальными клетками костного мозга. Является ростовым фактором пре-В- и пре-Т-лимфоцитов.
Ил-8 синтезируется моноцитами, макрофагами, фибробластами. Вызывает миграцию нейтрофилов и базофилов в очаг воспаления и их дегрануляцию, выделение супероксидного радикала. Стимулирует ангиогенез.
Ил-9 продуцируется гл. обр. Т-лимфоцитами. Стимулирует пролиферацию Т-лимфоцитов, активирует тучные клетки, усиливает эффекты эритропоэтина.
Ил-10 синтезируется Th2 а также цитотоксическими Т-лимфоцитами второго порядка и макрофагами. Стимулирует пролиферацию и дифференцировку В-лимфоцитов, подавляет синтез Ил-2 и ИФН-у клетками Thl, угнетает продукцию провоспалительных цитокинов.
Ил-11 продуцируется стромальными клетками костного мозга. Стимулирует деление и дифференцировку предшественников гемопоэза, колониеобразование мегакариоцитов, увеличивает количество тромбоцитов и эритроцитов в периферической крови. Угнетает продукцию провоспалительных цитокинов.
Ил-12 продуцируют моноциты, макрофаги и, в меньшей степени, В-лимфоциты и дендритные клетки. Стимулирует рост и дифференцировку Th (Th0 => Thl), Т-киллеров, NK. Индуцирует продукцию ИФН-g Т-лимфоцитами и NK, угнетает апоптоз Thl, синтез IgE. Вместе с ИЛ-4 регулирует баланс Thl и Th2.
Ил-13 синтезируется Th2. Стимулирует рост и дифференцировку В-лимфоцитов, подавляет функцию моноцитов/макрофагов, в частности секрецию провоспалительных цитокинов.
Ил-14 продуцируется в основном Т-лимфоцитами. Усиливает пролиферацию В-лимфоцитов и подавляет продукцию иммуноглобулинов.
Ил-15 вырабатывается моноцитами, эпителиоцитами и гладкомышечными клетками. По действию на Т-лимфоциты ИЛ-15 сходен с ИЛ-2, что объясняется способностью специфически связываться с ИЛ-2-рецепторами. Активирует NK и В-лимфоциты.
Ил-16 синтезируется эозинофилами и CD8+ Т-лимфоцитами. Является хемоаттрактантом для CD4+ лимфоцитов.
Ил-17 продуцируется активированными CD4 Т-лимфоцитами. Основными клетками-мишенями цитокина являются эпителиоциты, эндотелиоциты и фибробласты. Он усиливает выработку ИЛ-б, ИЛ-8, гранулоцитарного КСФ, простатландина Е2, увеличивает экспрессию ICAM-1, стимулирует активность фибробластов.
Ил-18 образуется активированными макрофагами, а также гепатоцитами. Стимулирует синтез Т-лимфоцитами ИФН-g, макрофагами – ИЛ-1, ИЛ 8 и ФНО. Кроме того, он активирует NK.
Факторы некроза опухоли
Различают: собственно фактор некроза опухоли (ФНО), или ФНОα лимфотоксины, или ФНОβ. ФНОα продуцируется макрофагами, а также тучными клетками и лимфоцитами. Он обусловливает развитие токсического шока и кахексии (старое название кахектин), индуцирует острофазные белки и стимулирует ангиогенез. Может индуцировать апоптоз. Способен вызывать геморрагический некроз ряда опухолей. ФНОα продуцируется Т- и В-лимфоцитами, обладает аналогичным действием.
Интерфероны
Интерфероны – гликопротеины, вырабатываемые клетками в ответ на вирусную инфекцию и другие стимулы. Блокируют репликацию вируса в других клетках и участвуют во взаимодействии между клетками иммунной системы. Различают две серологические группы интерферонов: I типа – ИНФ-α и -β и II типа – ИФН-γ.
Интерфероны I типа оказывают противовирусные и противоопухолевые эффекты, в то время как интерферон II типа регулирует специфический иммунный ответ и неспецифическую резистентность.
ИНФ-α и ИНФ-β отличаясь по структуре и клеткам-продуцентам, обладают практически одинаковым механизмом действия. В норме ИНФ-α продуцируется мононуклеарными фагоцитами (отсюда одно из названий – “лейкоцитарный ЛФН”), а ИНФ-β – фибробластами (“фибробластный ИФН”) . Под воздействием микроба секретируются многими клетками. Усиливают продукцию ИФН пирогеиное действие ИЛ-1 и понижение рН в межклеточной жидкости на фоне повышения температуры. Защитное действие ИНФ I типа реализуется посредством ингибирования репликации РНК или ДНК под воздействием олигоаденилат-синтетазы, которую продуцируют интерферон-содержащие клетки. ИНФ I типа, связываясь со здоровыми клетками, защищает их от вирусов.
Антивирусное действие ИНФ I типа может обусловливаться и тем, что он способен угнетать клеточную пролиферацию, препятствуя синтезу аминокислот, например триптофана. Этот механизм, а также способность индуцировать програмированную клеточную гибель некоторых опухолей лежат в основе противоопухолевого действия ИФН I типа. Кроме того, ИНФ I типа усиливает литическое действие нормальных киллеров на клетки-мишени, в том числе трансформированные клетки, индуцирует экспрессию антигенов МНС I и, наоборот, подавляет формирование тех же антигенов МНС II.
ИФН-γ (“иммунный ИФН”) продуцируется Т-лимфоцитами и NK. Стимулирует активность Т- и В-лимфоцитов, моноцитов/макрофагов и нейтрофилов. Усиливает экспрессию молекул МНС I, МНС II. Стимулирует дифференцировку ThO в Thl. Иммунный ИФН вместе со своим антогонистом ИЛ-4 поддерживает баланс Thl/Th2. Помимо этого, ИФН-γ регулирует апоптоз целого ряда нормальных, а также некоторых инфицированных и трансформированных клеток. Так, он индуцирует програмированную клеточную гибель активированных макрофагов, кератиноцитов, гепатоцитов, клеток костного мозга, эндотелиоцитов и подавляет апоптоз периферических моноцитов и герпес-инфицированных нейронов.
Колониестимулирующие факторы (КСФ)
Цитокины, регулирующих деление, дифференцировку костно-мозговых стволовых клеток и предшественников клеток крови. Кроме того, они могут стимулировать дифференцировку и функциональную активность некоторых клеток вне костного мозга.
- Гранулоцитарный КСФ (Г-КСФ) продуцируется в основном макрофагами, а также фибробластами. Стимулирует деление и дифференцировку стволовые клеток, в некоторой степени усиливает активность нейтрофилов и эозинофилов.
- Макрофагальный КСФ (М-КСФ) вырабатывается моноцитами, в меньшей степени эндотелиальными клетками и фибробластами. Активирует пролиферации предшественников макрофагов в костном мозге.
- Гранулоцитарно-макрофагальный КСФ (ГМ-КСФ) продуцируется макрофагами И Т-лимфоцитами, а также фибробластами и эндотелиоцитами. Стимулирует деление и дифференцировку предшественников гранулоцитов и макрофагов, активирует функцию макрофагов и гранулоцитов, пролиферацию Т-клеток. Участвует в стимуляции дифференцировки кроветворных предшестенников Е антигенпрезентирующие дендритные клетки.
Трансформирующий фактор роста-β (ТФР-β)
Наиболее изученный полифункциональный ростовой фактор, к которому относятся также фактор роста фибробластов, тромбоцитов, эндотелия, инсулиноподобный фактор роста, эпидермальный ростовой фактор и др. ТФР-β продуцируется многими клетками (основные продуценты – макрофаги), в том числе некоторыми опухолевыми клетками. ТФР-β – мощный деактивирующий фактор для моноцитов/макрофагов, существенно снижая их цитотоксическую и цитокин-продуцирующую активность, а также экспрессию на их поверхности молекул МНС.
В этом отношении он действует синергично с другими макрофаг-деактивирующими цитокинами (ИЛ-4, -10 и -13) . ТФР-β относят к преимущественно противовоспалительным цитокинам, благодаря его способности снижать продукцию нитросоединений, реакционно-способных радикалов и провоспалительных цитокинов клетками моноцитарно-макрофагального ряда. Однако в ряде случаев он способен оказывать и провоспалительные эффекты.
Большое значение имеет регуляция процессов программированной клеточной гибели нормальных и трансформированных клеток. ТФР-β угнетает апоптоз Thl, а вместе с ИЛ-2 ингибирует апоптоз Th2. Вероятно, ТФР-β, угнетая апоптоз клеток иммунной системы, играет важную роль в генерации клеток памяти. Избыточная активность этого и некоторых других ростовых факторов может приводить к гиперпролиферативным процессам, таким как гломерулонефрит, склерозированию кожи, циррозу печени и др., а также к прогрессирующему опухолевому росту. ТФР-β – один из медиаторов, обусловливающих иммуносупрессию при неопластических заболеваниях.
Хемокины
Основная функция хемокинов состоит в контроле клеточной миграции. Клетки, мигрирующие под влиянием хемокинов, следуют градиенту концентрации сигнала по направлению к его источнику. Одни хемокины контролируют клетки иммунной системы, направляя лимфоциты к лимфатическим узлам для того, чтобы те отслеживали проникающие в организм патогены за счёт взаимодействия с антиген-представляющими клетками. Это – гомеостатические хемокины, секретирующиеся постоянно и не нуждающиеся для этого в особых сигналах. Другая группа хемокинов участвует в развитие ткани, в частности, стимулируя ангиогенез, т.е. рост новых кровеносных сосудов, или направляя клетки в соответствующие ткани, где они дифференцируются. Наконец, воспалительные хемокины выделяются многими клетками в ответ на бактериальную инфекцию, вирусы или другие повреждающие агенты. Определённые хемокины могут инициировать иммунный ответ или стимулировать заживление раны. Хемокины секретируются активированными эндотелиальными и эпителиальными клетками, фибробластами, нейтрофилами и моноцитами и некоторыми другими клетками.
Цитокиновая сеть – саморегулирующаяся система, нарушение в которой приводит к избыточному или недостаточному синтезу определенных цитокинов, что в свою очередь может способствовать развитию разнообразных патологических процессов.